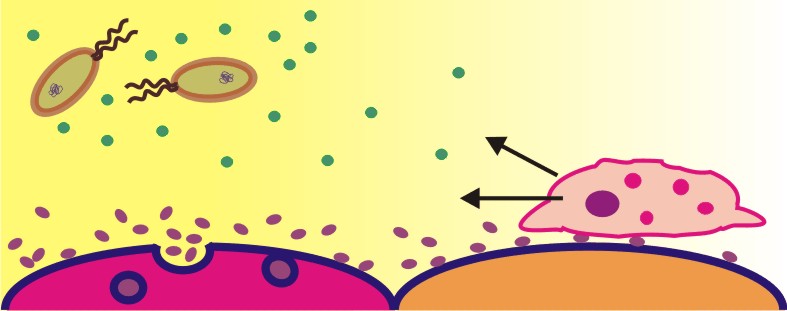
Was tun Zellen, wenn sie in Not geraten? Sie rufen um Hilfe – nicht mit akustischen Reizen, sondern mit chemischen Signalen. Fast alle Zellen des menschlichen Körpers können, wenn sie von Pathogenen oder durch Verletzungen gereizt werden, Signalproteine abgeben, die die Zellen des Immunsystems anlocken. Diese chemotaktischen Zytokine oder kurz Chemokine, diffundieren durch das Gewebe und bilden dabei ein Konzentrationsgefälle aus. Viele Arten von Immunzellen besitzen Rezeptoren, mit denen sie Chemokine erkennen können. Durch die Bindung eines Chemokins an den Rezeptor wird in der Zelle eine Signalkaskade ausgelöst, die dazu führt, dass die Zelle in Richtung steigender Chemokinkonzentration also hin zum gereizten oder entzündeten Gewebe, wandert. Im entzündeten Gewebe bewirken Chemokine und andere Entzündungsmediatoren, so dass die Immunzellen reaktive Sauerstoffspezies und Enzyme ausschütten, die mögliche bakterielle Eindringlinge abtöten sollen.
Auch wenn Chemokine nicht die einzigen Moleküle sind, die die Immunantwort steuern, ist das Chemokinsystem komplex: 50 verschiedene Chemokine konnten im Menschen identifiziert werden, dazu 20 Rezeptoren. Manche Chemokine binden nur an einen Rezeptor, andere an mehrere, genau wie manche Rezeptoren nur von einem Chemokin aktiviert werden können und andere von einer ganzen Reihe dieser Botenstoffe eingeschaltet werden. Dabei werden die Chemokine nach Anzahl und Position der Cysteinreste am N-Terminus in vier Gruppen (CC, CXC, C und CX3C) unterteilt. Die Funktion von Chemokinen ist nicht auf Entzündungs- reaktionen begrenzt. Sie regeln auch die Wanderung von Stammzellen zu ihrem Zielort in der Embryonal- entwicklung oder bei der Wundheilung, und sie rufen Immunzellen zu den Lymphknoten zurück, um das Immunsystem im Ruhezustand zu kontrollieren. [Allen, 2007]
Darüber hinaus spielen Chemokine eine Rolle bei verschiedenen Krankheiten, insbesondere bei chronischen Entzündungen und Autoimmunerkrankungen, wie rheumatoider Arthritis, Schuppenflechte, Asthma oder chronischen Darmentzündungen. Die Schwere der Krankheiten hängt dabei häufig mit der Zahl der (durch Chemokine rekrutierten) eingewanderten Leukozyten zusammen. Die Inhibition der Chemokin-Rezeptorwechselwirkung sollte diese Einwanderung stoppen und damit das entzündete Gewebe entlasten. Chemokininhibitoren sind daher interessante Kandidaten für neue pharmazeutische Wirkstoffe. [Wells, 2006]
Auch bei der Krebsentstehung spielen Chemokine eine Rolle. So regen sie die Neubildung von Blutgefäßen in Tumoren an und sicher deren Versorgung oder steuern die Wanderung von Metastasen, deren Zellen häufig Chemokinrezeptoren exprimieren. Ein weiterer Grund Chemokine zu inhibieren.
In unserer Arbeitsgruppe konzentrieren wir uns auf Interleukin-8 (IL-8) und Eotaxin als Entzündungsmediatoren und auf SDF-1α (stromal cell derived factor), ein Chemokin, das die Migration von Stammzellen (und Krebszellen) steuert.
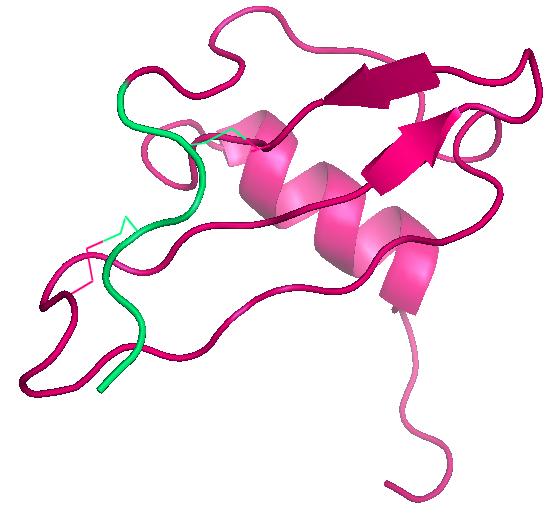
IL-8 (Abb. 2) gehört zu den inflammatorischen Chemokinen und bindet an die Rezeptoren CXCR1 und CXCR2. Diese beiden Rezeptoren werden in neutrophilen Granulozyten, aber auch in Monozyten exprimiert. Interleukin-8 kann in hohen Ausbeuten in E. coli exprimiert werden und ist damit auch hervorragend zur Entwicklung von Nachweisverfahren und proteinchemischen Methoden geeignet.
Weiterführende Literatur
Wells, T. N.; Power, C. A.; Shaw, J. P. und Proudfoot, A. E.; Trends Pharmacol. Sci. 2006, 27, 41-47.
Allen, S.; Crown, S. E. und Handel, T. M. Annu. Rev. Immunol. 2007, 25, 787–820,
Zlotnik, A. und Yoshie, O.; Immunity 2012, 36, 705-716.
